
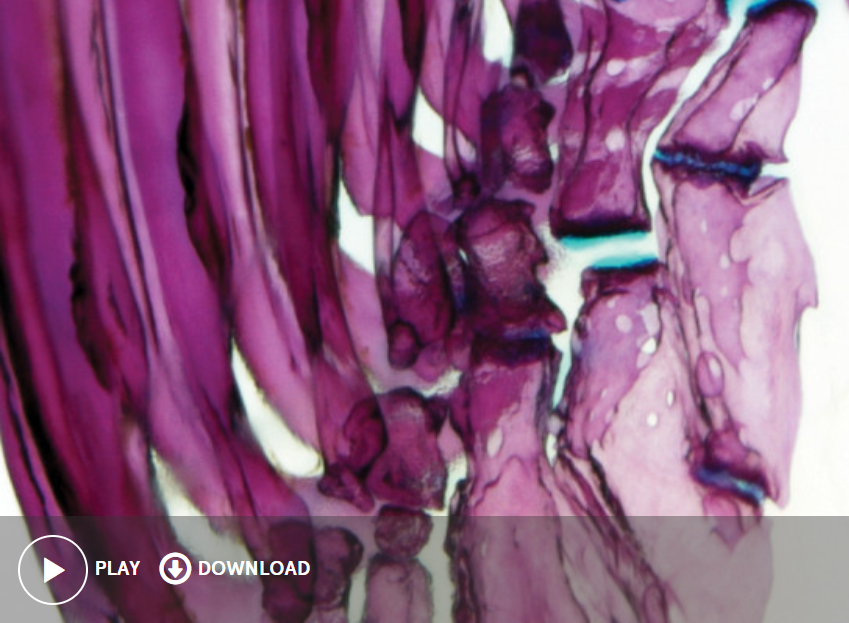
Flimbs and developmental potential
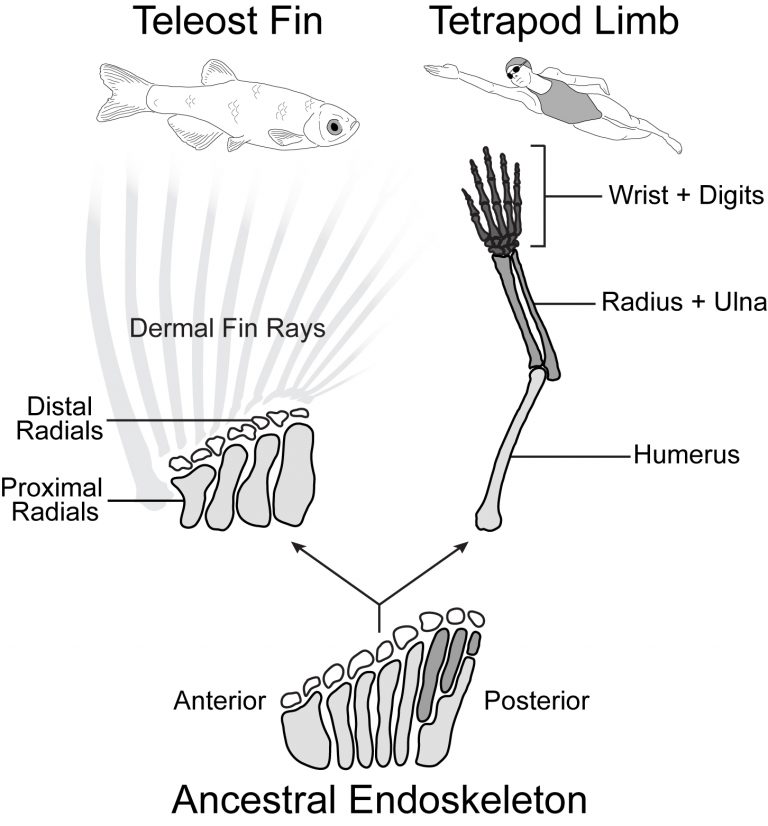
Although the forelimbs of tetrapods and the pectoral fins of teleost fishes have radically different morphologies, these divergent structures arose from a common appendage in the bony fish ancestor. From this ancestral appendage, elements were added to the proximal-distal (shoulder to fingertip) axis of the appendage in the tetrapod lineage, resulting in complex and elaborated limbs. In contrast, teleosts evolved a simplified pectoral fin through the loss of skeletal elements. The limb is divided into three regions along the proximal-distal axis: the proximal stylopod (humerus), the middle zeugopod (radius and ulna), and the distal autopod (wrist and hand). A more simple structure, the teleost pectoral fin is divided into two regions: the the proximal endoskeleton (radials) and the distal dermal skeleton (fin rays). How was the ancestral fin patterning program modified to produce complex, elaborated limbs in the tetrapod lineage and simplified pectoral fins in teleosts? Is the ability to form limbs unique to the tetrapod lineage, or was this ability latent in the bony fish ancestor?
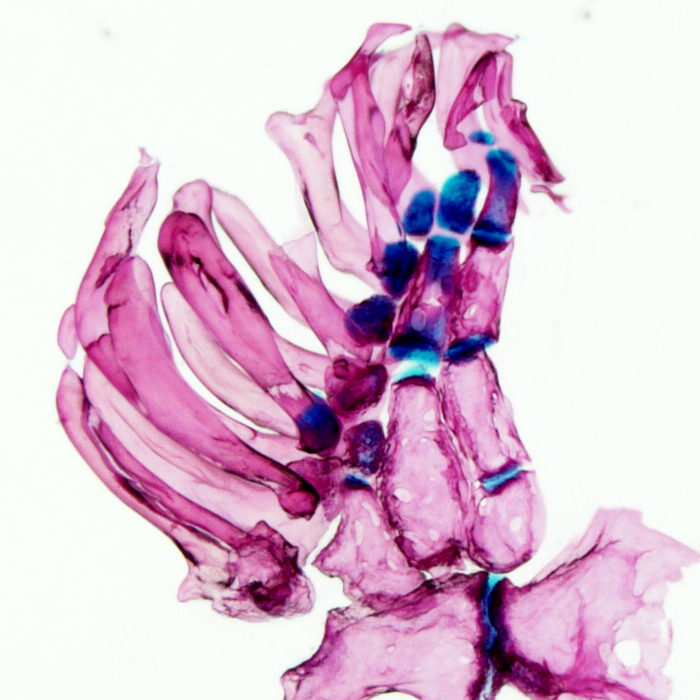
Using forward mutagenesis in zebrafish we recently discovered the mutant rephaim that forms novel bones along the proximal-distal axis of the fin endoskeleton in a limb-like manner. We found that Hox11 genes are required to form the new bones and that hoxa11b is unregulated in the rephaim mutant. Interestingly, Hox11 genes are also required for the normal development of the middle limb region, the zeugopod. These findings suggest that the ability to specify a middle appendage region was already present in the bony fish ancestor, and this potential was realized in tetrapods but remains latent in teleosts. Considering this result with another recent discovery that both fins and limbs require Hox13 to form their most distal regions, a paradigm is emerging that proximal-distal patterning mechanisms are conserved between fins and limbs and likely arose in the bony fish common ancestor.
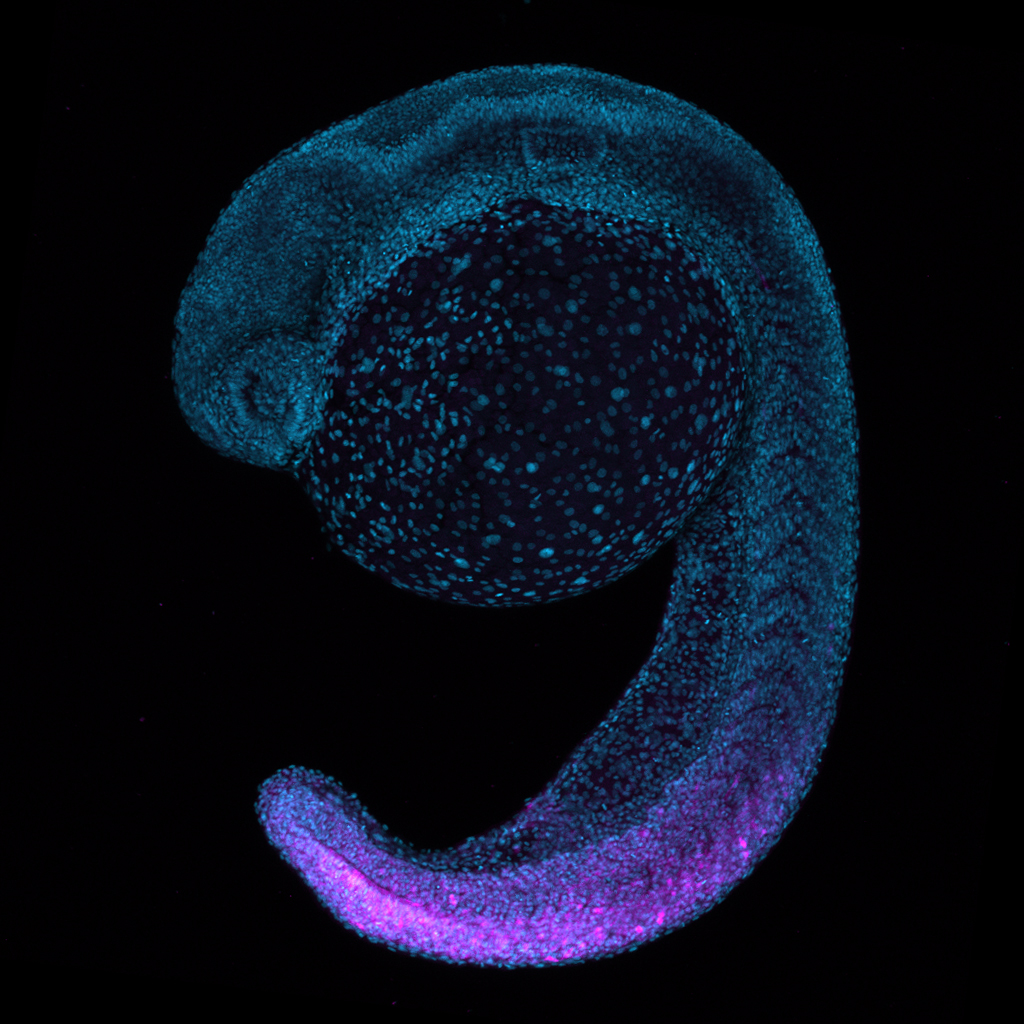
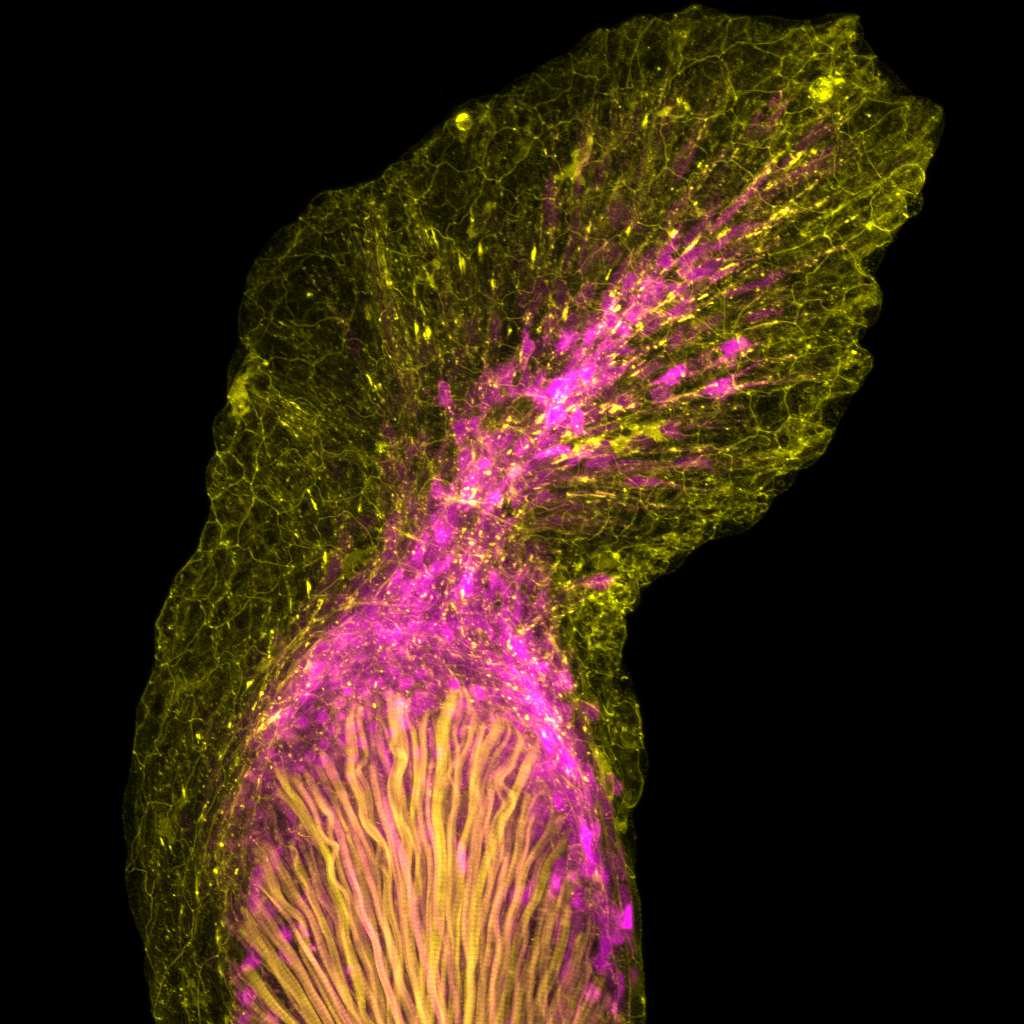

Given the divergent anatomies of fins and limbs, it is difficult to assign one-to- one homology relationships between fin and limb elements. However, developmental genetic studies have revealed genes and patterning programs that regulate specific regions of limbs, and studying these candidates in fins holds promise to reveal deep homology relationships between appendage regions. The aforementioned studies suggest that similar mechanisms underlie the formation the middle and distal regions of fins and limbs. Now, I am using knock-in tools and new mutants with proximal appendage phenotypes to investigate proximal appendage and shoulder patterning in fins and limbs.